

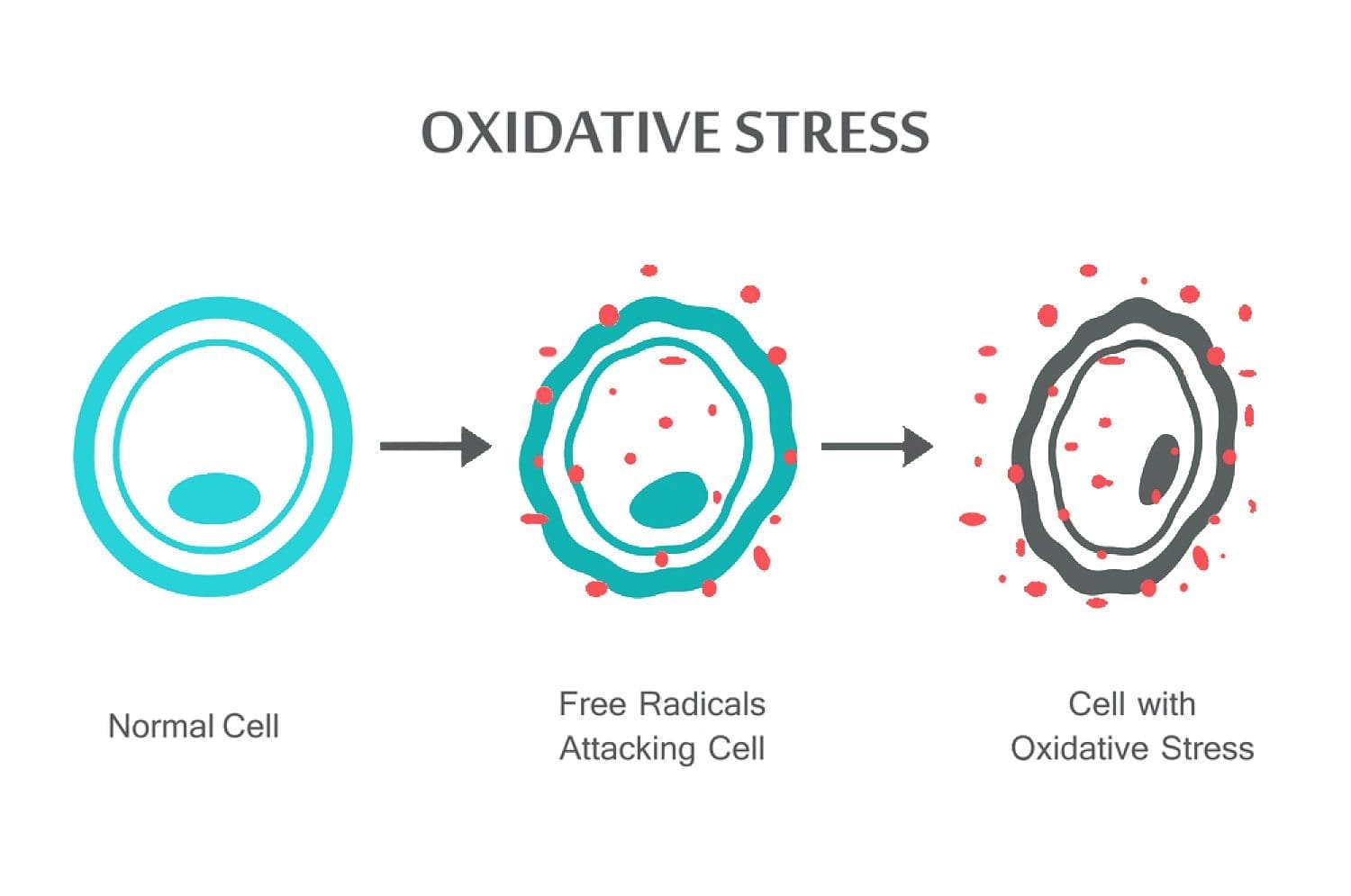
Ilmu pengetahuan berbasis Chiropractor Dr. Alexander Jimenez lihatlah stres oksidatif, apa itu, bagaimana hal itu mempengaruhi tubuh dan antioksidan pertahanan untuk memperbaiki situasi.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD, 2 dan Omer Kalayci, MD1
Abstrak: Spesies oksigen reaktif (ROS) diproduksi oleh organisme hidup sebagai hasil metabolisme sel normal dan faktor lingkungan, seperti polutan udara atau asap rokok. ROS adalah molekul yang sangat reaktif dan dapat merusak struktur sel seperti karbohidrat, asam nukleat, lipid, dan protein dan mengubah fungsinya. Pergeseran keseimbangan antara oksidan dan antioksidan yang mendukung oksidan disebut "stres oksidatif." Peraturan tentang pengurangan dan pengoksidasi (redoks) sangat penting untuk kelangsungan hidup sel, aktivasi, proliferasi, dan fungsi organ. Organisme aerobik memiliki sistem antioksidan terintegrasi, yang mencakup antioksidan enzimatik dan non-enzimatik yang biasanya efektif dalam menghambat efek berbahaya ROS. Namun, dalam kondisi patologis, sistem antioksidan bisa terbebani. Stres oksidatif berkontribusi terhadap banyak kondisi patologis dan penyakit, termasuk kanker, gangguan neurologis, aterosklerosis, hipertensi, iskemia / perfusi, diabetes, sindrom gangguan pernafasan akut, fibrosis paru idiopatik, penyakit paru obstruktif kronik, dan asma. Dalam tinjauan ini, kami merangkum sistem oksidan dan antioksidan seluler dan mendiskusikan efek seluler dan mekanisme stres oksidatif.
Kata kunci: antioksidan, oksidan, stres oksidatif, spesies oksigen reaktif, redoks
(Jurnal WAO 2012; 5: 9-19)
Spesies oksigen reaktif (ROS) diproduksi oleh organisme hidup sebagai hasil metabolisme sel normal. Pada konsentrasi rendah hingga sedang, mereka berfungsi dalam proses sel fisiologis, tetapi pada konsentrasi tinggi, mereka menghasilkan modifikasi yang merugikan pada komponen sel, seperti lipid, protein, dan DNA.1–6 Pergeseran keseimbangan antara oksidan / antioksidan yang mendukung oksidan disebut "stres oksidatif". Stres oksidatif berkontribusi pada banyak kondisi patologis, termasuk kanker, gangguan neurologis, 7-10 aterosklerosis, hipertensi, iskemia / perfusi, diabetes 11-14, sindrom gangguan pernapasan akut, fibrosis paru idiopatik, penyakit paru obstruktif kronik, 15 dan asma.16– 21 Organisme aerobik memiliki sistem antioksidan terintegrasi, yang mencakup antioksidan enzimatik dan nonenzimatik yang biasanya efektif dalam memblokir efek berbahaya dari ROS. Namun, dalam kondisi patologis, sistem antioksidan bisa kewalahan. Dalam ulasan ini, kami merangkum oksidan seluler dan sistem antioksidan dan regulasi keadaan reduksi dan oksidasi (redoks) dalam kondisi kesehatan dan penyakit.
Daftar Isi
OXIDANTS
Sumber Endogen dari ROS
ROS diproduksi dari molekul oksigen sebagai hasil metabolisme sel normal. ROS dapat dibagi menjadi kelompok 2: radikal bebas dan nonradis. Molekul yang mengandung satu atau lebih elektron tak berpasangan dan dengan demikian memberi reaktivitas terhadap molekul disebut radikal bebas. Bila radikal bebas 2 membagikan elektron mereka yang tidak berpasangan, bentuk non-statis telah dibuat. ROS utama 3 yang bermakna fisiologis adalah anion superoksida (O22.), Radikal hidroksil (OH), dan hydro-gen peroxide (H2O2). ROS dirangkum dalam Tabel 1.
 Anion superoksida dibentuk dengan penambahan 1 elektron ke molekul oksigen.22 Proses ini dimediasi oleh nikotin adenin dinukleotida fosfat [NAD (P) H] oksidase atau xantin oksidase atau oleh sistem transportasi elektron mitokondria. Situs utama untuk memproduksi anion superoksida adalah mitokondria, mesin sel untuk menghasilkan adenosin trifosfat. Biasanya, elektron ditransfer melalui rantai transpor elektron mitokondria untuk reduksi oksigen ke air, tetapi sekitar 1 hingga 3% dari semua elektron bocor dari sistem dan menghasilkan superoksida. NAD (P) H oksidase ditemukan dalam leukosit polimorfonuklear, monosit, dan makrofag. Setelah fagositosis, sel-sel ini menghasilkan ledakan superoksida yang menyebabkan aktivitas bakterisidal. Superoksida diubah menjadi hidrogen peroksida melalui aksi superoksida dismutase (SODs, EC 1.15.1.1). Hidrogen peroksida dengan mudah berdifusi melintasi membran plasma. Hidrogen peroksida juga diproduksi oleh xantin oksidase, asam amino oksidase, dan NAD (P) H oksidase 23,24 dan dalam peroksisom dengan konsumsi oksigen molekuler dalam reaksi metabolik. Dalam rangkaian reaksi yang disebut reaksi Haber – Weiss dan Fenton, H2O2 dapat terurai menjadi OH2 dengan adanya logam transmisi seperti Fe21 atau Cu21.25
Anion superoksida dibentuk dengan penambahan 1 elektron ke molekul oksigen.22 Proses ini dimediasi oleh nikotin adenin dinukleotida fosfat [NAD (P) H] oksidase atau xantin oksidase atau oleh sistem transportasi elektron mitokondria. Situs utama untuk memproduksi anion superoksida adalah mitokondria, mesin sel untuk menghasilkan adenosin trifosfat. Biasanya, elektron ditransfer melalui rantai transpor elektron mitokondria untuk reduksi oksigen ke air, tetapi sekitar 1 hingga 3% dari semua elektron bocor dari sistem dan menghasilkan superoksida. NAD (P) H oksidase ditemukan dalam leukosit polimorfonuklear, monosit, dan makrofag. Setelah fagositosis, sel-sel ini menghasilkan ledakan superoksida yang menyebabkan aktivitas bakterisidal. Superoksida diubah menjadi hidrogen peroksida melalui aksi superoksida dismutase (SODs, EC 1.15.1.1). Hidrogen peroksida dengan mudah berdifusi melintasi membran plasma. Hidrogen peroksida juga diproduksi oleh xantin oksidase, asam amino oksidase, dan NAD (P) H oksidase 23,24 dan dalam peroksisom dengan konsumsi oksigen molekuler dalam reaksi metabolik. Dalam rangkaian reaksi yang disebut reaksi Haber – Weiss dan Fenton, H2O2 dapat terurai menjadi OH2 dengan adanya logam transmisi seperti Fe21 atau Cu21.25
Fe31 + .O2? Fe2 + O2 Haber Weiss
Fe2 + H2O2? Fe3 + OH + .OH Reaksi Fenton
O 2 sendiri juga dapat bereaksi dengan H2 O2 dan menghasilkan OH .26,27 Radikal hidroksil merupakan ROS yang paling reaktif dan dapat merusak protein, lipid, serta karbohidrat dan DNA. Ia juga dapat memulai peroksidasi lipid dengan mengambil elektron dari asam lemak tak jenuh ganda.
Enzim granulocytic selanjutnya memperluas reaktivitas H2O2 melalui eosinophil peroxidase dan myeloperoxidase (MPO). Pada neutrofil aktif, H2O2 dikonsumsi oleh MPO. Dengan adanya ion klorida, H2O2 diubah menjadi asam hipoklorida (HOCl). HOCl sangat oksidatif dan berperan penting dalam membunuh patogen di jalan napas.28 Namun, HOCl juga dapat bereaksi dengan DNA dan menginduksi interaksi protein-DNA dan menghasilkan produk oksidasi pirimidin dan menambahkan klorida ke basis DNA.29,30 Eosinophil peroxidase dan MPO juga berkontribusi pada stres oksidatif dengan modifikasi protein dengan halogenasi, nitrasi, dan ikatan silang protein melalui radikal tirosil. 31-33
Radikal radikal lain yang berasal dari oksigen adalah radikal peroksil (ROO $). Bentuk paling sederhana dari radikal ini adalah radikal hidro- peroksil (HOO $) dan memiliki peran dalam peroksidasi asam lemak. Radikal bebas dapat memicu reaksi rantai peroksidasi lipid dengan mengabstraksikan atom hidrogen dari karbon metilen rantai samping. Radikal lipida kemudian bereaksi dengan oksigen untuk menghasilkan radikal peroksil. Radikal peroksil memulai reaksi berantai dan mengubah asam lemak tak jenuh ganda menjadi hydroperoxides lipid. Hydroperoxides lipida sangat tidak stabil dan mudah terurai menjadi produk sekunder, seperti aldehida (seperti 4-hydroxy-2,3-nonenal) dan malondialdehydes (MDA). Isoprostan adalah kelompok produk peroksidasi lipid lain yang dihasilkan melalui peroksidasi asam arakidonat dan juga ditemukan meningkat pada kondensat plasma dan nafas penderita asma.34,35 Peroksidasi lipid mengganggu integritas membran sel dan menyebabkan penataan kembali struktur membran. .
Hidrogen peroksida, radikal superoksida, glutathione teroksidasi (GSSG), MDA, isoprostan, karbonil, dan nitrotyrosin dapat dengan mudah diukur dari sampel lavage plasma, darah, atau bronchoalveolar sebagai biomarker oksidasi dengan uji standar.
Sumber Eksim Oksidan
Asap rokok
Asap rokok mengandung banyak oksidan dan radikal bebas dan senyawa organik, seperti superoksida dan oksida nitrat. 36 Selain itu, menghirup asap rokok ke paru-paru juga mengaktifkan beberapa mekanisme endogen, seperti akumulasi neutrofil dan makrofag, yang selanjutnya meningkatkan cedera oksidan. .
Paparan Ozon
Paparan ozon dapat menyebabkan peroksidasi lipid dan menyebabkan masuknya neutrofil ke epitel saluran napas. Paparan jangka pendek terhadap ozon juga menyebabkan pelepasan mediator inflamasi, seperti MPO, protein kationik eosinofilik dan juga dehidrogenase laktat dan albumin.37 Bahkan pada subyek sehat, paparan ozon menyebabkan pengurangan fungsi paru.38 Cho et al39 telah menunjukkan bahwa partikulat (campuran partikel padat dan tetesan cairan yang tersuspensi di udara) mengkatalisis pengurangan oksigen.
Hyperoxia
Hyperoxia mengacu pada kondisi tingkat oksigen yang lebih tinggi daripada tekanan parsial oksigen normal di paru-paru atau jaringan tubuh lainnya. Ini menyebabkan produksi yang lebih besar dari spesies oksigen dan nitrogen reaktif. 40,41
Radiasi pengion
Radiasi pengion, dengan adanya O2, mengubah radikal hidroksil, superoksida, dan radikal organik menjadi hidrogen peroksida dan hidroperoksida organik. Spesies hidroperoksida ini bereaksi dengan ion logam aktif redoks, seperti Fe dan Cu, melalui reaksi Fenton dan dengan demikian menginduksi stres oksidatif. 42,43 Narayanan et al44 menunjukkan bahwa fibroblas yang terpapar partikel alfa memiliki peningkatan yang signifikan pada 2 O2 intraselular. dan produksi H2O2 melalui oksidase NADPH-oksida kompleks. 44 Molekul-molekul transduksi sinyal, seperti Xenum Xenum dan 1 yang diatur regulasi ekstraselular (ERK2 / 1), c-Jun N-terminal kinase (JNK), dan p2, dan faktor transkripsi, seperti protein pengaktif-38 (AP-1), faktor nuklir-kB (NF-kB), dan p1, diaktifkan, yang menghasilkan ekspresi gen terkait respons radiasi.53-45 Ultraviolet A (UVA) memicu foton reaksi oksidatif dengan eksitasi fotosensitizer endogen, seperti porfirin, oksidase NADPH, dan riboflavin. 50-Oxo-8- dihydroguanine (7,8-oxoGua) adalah produk oksidasi DNA mediasi utama UVA yang dibentuk oleh oksidasi oksidan OH, oksida 8-elektron, dan oksigen singlet yang terutama bereaksi dengan guanine.1 Pembentukan kation radikal guanin dalam DNA terisolasi telah terbukti efisien terjadi melalui efek langsung dari radiasi pengion. 51 Setelah terpapar radiasi pengion, tingkat intraselular glutathione (GSH) menurun untuk jangka pendek namun kemudian meningkat lagi.52,53
Ion Logam Berat
Ion logam berat, seperti besi, tembaga, kadmium, merkuri, nikel, timbal, dan arsenik, dapat menyebabkan pembentukan radikal reaktif dan menyebabkan kerusakan sel akibat penipisan aktivitas enzim melalui peroksidasi dan reaksi lipid dengan protein nuklir dan DNA.55
Salah satu mekanisme yang paling penting dari generasi radikal bebas yang dimediasi logam adalah melalui reaksi tipe Fenton. Ion superoksida dan hidrogen peroksida dapat berinteraksi dengan logam transisi, seperti besi dan tembaga, melalui katalis reaksi Haber-Weiss / Fenton yang dikatalisis logam untuk membentuk radikal OH.
Metal31 1 $ O2 / XXUMX 21 O1 Haber Weiss Metal2 21 H1 O2 / XXUMX 2 OH 31 1 $ OH Reaksi Fenton
Selain mekanisme jenis Fenton dan Haber-Weiss, ion logam tertentu dapat bereaksi langsung dengan molekul seluler untuk menghasilkan radikal bebas, seperti radikal tiol, atau menginduksi jalur sinyal sel. Radikal ini juga dapat bereaksi dengan molekul tiol lainnya untuk menghasilkan O22 .. O22. diubah menjadi H2O2, yang menyebabkan penambahan radikal oksigen tambahan. Beberapa logam, seperti arsenit, menginduksi pembentukan ROS secara tidak langsung dengan aktivasi sistem produksi radikal di sel. 56
Arsenik adalah unsur yang sangat beracun yang menghasilkan berbagai ROS, termasuk superoksida (O2 2), oksigen tunggal (1O2), radikal peroksil (ROO), oksida nitrat (NO), hidrogen peroksida (H2O2), dan radikal peroksil dimetilarsinat [ CH3) 2AsOO] .57-59 Senyawa arsenik (III) dapat menghambat enzim antioksidan, terutama enzim yang bergantung pada GSH, seperti glutathione-S-transferases (GSTs), glutathione peroxidase (GSH-Px), dan GSH reduktase, melalui ikatan - kepada kelompok sulfhydryl (-SH) mereka. 60,61
Timbal meningkatkan peroksidasi lipid.62 Penurunan aktivitas enzim SOD dan GPX otak yang signifikan terjadi setelah paparan timbal. 63,64 Penggantian seng, yang berfungsi sebagai kofaktor untuk banyak enzim oleh timbal, menyebabkan inaktivasi enzim tersebut. Paparan timbal dapat menyebabkan penghambatan GST dengan mempengaruhi tiol jaringan.
ROS yang dihasilkan oleh reaksi katalis logam dapat membentuk basis DNA. Tiga substitusi dasar, G / C, G / T, dan C / T, dapat terjadi sebagai akibat kerusakan oksidatif oleh ion logam, seperti Fe21, Cu21, dan Ni21. Reid et al65 menunjukkan bahwa G / C didominasi oleh Fe21 sedangkan substitusi C / T oleh Cu21 dan Ni21.
Antioksidan
Tubuh manusia dilengkapi dengan berbagai antioksidan yang berfungsi untuk mengimbangi efek oksidan. Untuk semua tujuan praktis, ini dapat dibagi menjadi kategori 2: enzimatik (Tabel 2) dan nonenzymatic (Tabel 3).



Enzimatik Antioksidan
Antioksidan enzimatik utama paru-paru adalah SOD (EC 1.15.1.11), katalase (EC 1.11.1.6), dan GSH-Px (EC 1.11.1.9). Selain enzim utama ini, antioksidan lainnya, termasuk heme oxygenase-1 (EC 1.14.99.3), dan protein redoks, seperti thioredoxins (TRXs, EC 1.8.4.10), peroxiredoxins (PRXs, EC 1.11.1.15), dan glutaredoxins, juga ditemukan pada memainkan peran penting dalam pertahanan antioksidan paru.
Karena superoksida adalah ROS utama yang dihasilkan dari berbagai sumber, penurunannya oleh SOD sangat penting untuk setiap sel. Semua bentuk 3 dari SOD, yaitu CuZn-SOD, Mn-SOD, dan EC-SOD, banyak diekspresikan di paru-paru manusia. Mn-SOD dilokalisasi dalam matriks mitokondria. EC-SOD terutama terlokalisasi dalam matriks ekstraselular, terutama di daerah-daerah yang mengandung serat kolagen tipe I yang tinggi dan sekitar pembuluh darah paru dan sistemik. Ini juga telah terdeteksi di epitel bronkial, epitel alveolar, dan makrofag alveolar.66,67 Secara keseluruhan, CuZn-SOD dan Mn-SOD pada umumnya dianggap bertindak sebagai pemulung massal radikal superoksida. Tingkat EC-SOD yang relatif tinggi di paru-paru dengan pengikatnya yang spesifik terhadap komponen matriks ekstraselular dapat mewakili komponen dasar proteksi matriks paru.68
H2O2 yang diproduksi oleh aksi SOD atau tindakan oksidase, seperti xanthine oxidase, dikurangi menjadi air dengan katalase dan GSH-Px. Katalase ada sebagai tetra- mer yang terdiri dari monomer identik 4, yang masing-masing mengandung gugus heme di tempat yang aktif. Degradasi H2O2 dilakukan melalui konversi antara konformasi 2 dengan katalase-ferricatalase (besi yang dikoordinasikan dengan air) dan senyawa I (besi yang dikomplekskan dengan atom oksigen). Katalase juga mengikat NADPH sebagai padanan pereduksi untuk mencegah inaktivasi oksidatif enzim (pembentukan senyawa II) oleh H2O2 karena direduksi menjadi air.69
Enzim dalam siklus redoks bertanggung jawab untuk pengurangan H2O2 dan hydroperoxides lipid (dihasilkan sebagai hasil peroksidasi lipid membran) termasuk GSH-Pxs.70 GSH-Pxs adalah keluarga enzim tetramer yang mengandung selenosistein asam amino unik di dalam situs aktif dan menggunakan tiol berberat molekul rendah, seperti GSH, untuk mengurangi H2O2 dan peroksida lipida ke alkohol yang sesuai. Empat GSH-Pxs telah dijelaskan, dikodekan oleh gen yang berbeda: GSH-Px-1 (GSH-Px) seluler ada di mana-mana dan mengurangi H2O2 dan peroksida asam lemak, namun tidak mengoksidasi lipid peroksil.71 Lipid yang diferensikan dikurangi dengan GSH yang terikat membran -Px-4 (phospholipid hydroperoxide GSH-Px), yang dapat menggunakan beberapa tiol berberat molekul rendah yang berbeda seperti mengurangi padanannya. GSH-Px-2 (gastrointestinal GSH-Px) dilokalisasi di sel epitel gastrointestinal di mana berfungsi untuk mengurangi peroksida diet.72 GSH-Px-3 (GSH-Px ekstraselular) adalah satu-satunya anggota keluarga GSH-Px yang tinggal di kompartemen ekstraselular dan diyakini sebagai salah satu enzim antioksidan ekstraselular terpenting pada mamalia. Dari jumlah tersebut, GSH-Px ekstraselular paling banyak diteliti di paru-paru manusia.73
Selain itu, pembuangan H2O2 terkait erat dengan beberapa enzim yang mengandung tiol, yaitu TRXs (TRX1 dan TRX2), reduktase tioredoksin (EC 1.8.1.9) (TRRs), PRXs (yang merupakan peroksidase tioredoksin), dan glutaredoksin.74
Dua TRXs dan TRRs telah dicirikan pada sel manusia, ada di sitosol dan mitokondria. Di paru-paru, TRX dan TRR diekspresikan dalam epitel bronkial dan alveolar dan makrofag. Enam PRX yang berbeda telah ditemukan di sel manusia, berbeda dalam kompartementalisasi ultrastruktural. Studi eksperimental telah mengungkapkan pentingnya PRX VI dalam melindungi epitel alveolar. Paru manusia mengungkapkan semua PRXs di epitel bronkial, epitel alveolar, dan makrofag.75 PRX V baru-baru ini ditemukan berfungsi sebagai peroxynitrite reductase, 76 yang berarti berfungsi sebagai senyawa pelindung potensial dalam pengembangan cedera paru yang dimediasi ROS. .77
Yang umum untuk antioksidan ini adalah kebutuhan NADPH sebagai ekuivalensi pereduksi. NADPH mempertahankan katalase dalam bentuk aktif dan digunakan sebagai kofaktor oleh TRX dan GSH reductase (EC 1.6.4.2), yang mengubah GSSG menjadi GSH, co-substrat untuk GSH-Pxs. NADPH intraselular, pada gilirannya, dihasilkan oleh pengurangan NADP1 oleh glukosa-6-fosfat dehidrogenase, enzim pengikatan pertama dan laju batas dari jalur fosfat pen- tose, selama konversi glukosa-6-fosfat menjadi 6-phosphogluconolactone. Dengan menghasilkan NADPH, glukosa-6-phosphate dehydrogenase adalah penentu penting kapasitas buffer GSH sitosolik (GSH / GSSG) dan oleh karena itu dapat dianggap sebagai enzim antioksidan penting dan penting. 78,79
GST (EC 2.5.1.18), keluarga enzim antioksidan lainnya, melumpuhkan metabolit sekunder, seperti aldehida tak jenuh, epoksida, dan hidroperoksida. Tiga keluarga utama GST telah dijelaskan: GST sitosolik, GST mitokondria, 80,81 dan GST mikrosomal yang terkait dengan membran yang memiliki peran dalam metabolisme eikosanoid dan GSH. 82 Tujuh kelas GST sitosolik diidentifikasi pada mamalia, yang ditunjuk Alpha, Mu, Pi, Sigma, Theta, Omega, dan Zeta.83-86 Selama kondisi yang tidak tertekan, kelas Mu dan Pi GST berinteraksi dengan kinase Ask1 dan JNK, masing-masing, dan menghambat kinase ini.87-89 Telah ditunjukkan bahwa GSTP1 terdisosiasi dari JNK di respon terhadap stres oksidatif.89 GSTP1 juga berinteraksi secara fisik dengan PRX VI dan mengarah pada pemulihan aktivitas enzim PRX melalui glutathionylation dari protein teroksidasi.90
Antioksidan nonenzymatik
Antioksidan nonenzimatik termasuk senyawa dengan berat molekul rendah, seperti vitamin (vitamin C dan E), b-karoten, asam urat, dan GSH, sebuah tripeptida (Lg-glutamyl-L-cysteinyl-L-glineine) yang terdiri dari tiol ( sulfhidril).
Vitamin C (Asam askorbat)
Vitamin C yang larut dalam air (asam askorbat) menyediakan kapasitas antioksidan fase berair intraselular dan ekstraselular terutama dengan mengais oksigen bebas oksigen. Ini mengubah vitamin E radikal bebas kembali ke vitamin E. Tingkat plasmanya telah terbukti menurun seiring bertambahnya usia.91,92
Vitamin E (a-tokoferol)
Vitamin E yang larut dalam lipid terkonsentrasi di situs interior hidrofobik membran sel dan merupakan pertahanan utama terhadap cedera membran akibat oksidan. Vitamin E menyumbangkan elektron ke radikal peroksil, yang diproduksi selama peroksidasi lipid. a-Tokoferol adalah bentuk paling aktif vitamin E dan antioksidan utama yang terikat pada sel. Vitamin E memicu apoptosis sel kanker dan menghambat formasi radikal bebas.93
Glutathione
GSH sangat melimpah di semua kompartemen sel dan merupakan antioksidan utama yang dapat larut. Rasio GSH / GSSG merupakan penentu utama stres oksidatif. GSH menunjukkan efek antioksidannya dalam beberapa cara. 94 Ini mendetoksifikasi peroksida hidrogen peroksida dan lipid melalui aksi GSH-Px. GSH menyumbangkan elektronnya ke H2O2 untuk menguranginya menjadi H2O dan O2. GSSG kembali diturunkan menjadi GSH oleh reduktase GSH yang menggunakan NAD (P) H sebagai donor elektron. GSH-Pxs juga penting untuk melindungi selaput sel dari peroksidasi lipid. Mengurangi glutathione menyumbangkan proton pada lipida membran dan melindungi mereka dari serangan oksidan.95
GSH adalah kofaktor untuk beberapa enzim detoksifikasi, seperti GSH-Px dan transferase. Ini memiliki peran dalam mengubah vitamin C dan E kembali ke bentuk aktifnya. GSH melindungi sel-sel melawan apoptosis dengan berinteraksi dengan jalur pensinyalan proapoptosis dan antiapoptotik. 94 Ini juga mengatur dan mengaktifkan beberapa faktor transkripsi, seperti AP-1, NF-kB, dan Sp-1.
Karotenoid (b-karoten)
Karotenoid adalah pigmen yang ditemukan pada tumbuhan. Terutama, b-karoten telah ditemukan bereaksi dengan radikal peroksil (ROO), hidroksil (OH), dan superoksida (O22.) .96 Karotenoid menunjukkan efek antioksidannya pada tekanan parsial oksigen rendah namun memiliki efek pro-oksidan pada oksigen yang lebih tinggi. konsentrasi.97 Baik karotenoid dan asam retinoat (RAs) mampu mengatur faktor transkripsi.98 b-Carotene menghambat aktivasi NF-kB yang diinduksi oleh oksidan dan interleukin (IL) -6 dan faktor nekrosis tumor - sebuah produksi. Karotenoid juga mempengaruhi apoptosis sel. Efek antiproliferatif RA telah ditunjukkan pada beberapa penelitian. Efek RA ini dimediasi terutama oleh reseptor asam retinoat dan bervariasi di antara jenis sel. Pada sel karsinoma mammae, reseptor asam retinoat ditunjukkan untuk memicu penghambatan pertumbuhan dengan menginduksi penangkapan sel siklus, apoptosis, atau keduanya. 99,100
PENGARUH STRES OKSIDATIF: MEKANISME GENETIK, FISIOLOGI, & BIOKIMIA
Stres oksidatif terjadi ketika keseimbangan antara antioksidan dan ROS terganggu karena penipisan antioksidan atau akumulasi ROS. Ketika stres oksidatif terjadi, sel mencoba untuk melawan efek oksidan dan mengembalikan keseimbangan redoks dengan mengaktifkan atau membungkam gen yang mengkode enzim defensif, faktor tranfiksi, dan protein struktural. Rasio 101,102 antara glutathione teroksidasi dan glukosa (2GSH / GSSG) adalah satu faktor penentu stres oksidatif yang penting dalam tubuh. Produksi ROS yang lebih tinggi dalam tubuh dapat mengubah struktur DNA, menghasilkan modifikasi protein dan lipid, pengaktifan beberapa faktor transkripsi yang disebabkan oleh stres, dan produksi sitokin pro-inflamasi dan anti-inflamasi.
Efek Stres Oksidatif Pada DNA
ROS dapat menyebabkan modifikasi DNA dengan beberapa cara, yang melibatkan degradasi basa, pemecahan DNA tunggal atau double-stranded, purin, pirimidin atau modifikasi terikat gula, mutasi, penghapusan atau translokasi, dan hubungan silang dengan protein. Sebagian besar modifikasi DNA (Gambar 1) sangat relevan untuk karsinogenesis, penuaan, dan penyakit neurodegeneratif, kardiovaskular, dan autoimun. Asap tembakau, logam redoks, dan logam nonredoks, seperti besi, kadmium, krom, dan arsenik, juga terlibat dalam karsinogenesis dan penuaan dengan menghasilkan radikal bebas atau ikatan dengan kelompok tiol. Pembentukan 8-OH-G adalah kerusakan DNA yang paling dikenal yang terjadi melalui stres oksidatif dan merupakan biomarker potensial untuk karsinogenesis.

Area promoter gen mengandung sekuens konsensus untuk faktor transkripsi. Situs pengikatan faktor transkripsi ini mengandung urutan kaya GC yang rentan terhadap serangan oksidan. Pembentukan DNA 8-OH-G di tempat pengikatan faktor transkripsi dapat memodifikasi pengikatan faktor transkripsi dan dengan demikian mengubah ekspresi gen terkait seperti yang telah ditunjukkan untuk urutan target AP-1 dan Sp-1.103 Selain 8-OH-G, 8,59 -cyclo-29 -deoxyadenosine (cyclo-dA) juga telah terbukti menghambat transkripsi dari gen reporter dalam sistem sel jika berada dalam kotak TATA.104 Protein pengikat TATA memulai transkripsi dengan mengubah lentur DNA. Pengikatan protein pengikatan TATA dapat terganggu oleh adanya siklo-dA.
Stres oksidatif menyebabkan ketidakstabilan daerah mikrosatelit (daerah persendian pendek). Redoks ion logam aktif, radikal hidroksil meningkatkan ketidakstabilan mikrosatelit.105 Meskipun pemecahan DNA untai tunggal yang disebabkan oleh luka oksidan dapat ditoleransi dengan mudah oleh sel-sel, kerusakan DNA untai ganda yang disebabkan oleh radiasi pengion dapat menjadi ancaman signifikan bagi kelangsungan hidup sel. 106
Metilasi di pulau CpG dalam DNA adalah mekanisme epigenetik penting yang dapat menyebabkan pembungkaman gen. Oksidasi 5-MeCyt menjadi 5-hydroxymethyl uracil (5-OHMeUra) dapat terjadi melalui reaksi deaminasi / oksidasi intermediet timin atau 5-hydroxymethyl cytosine.107 Selain ekspresi gen modulasi, metilasi DNA juga tampaknya mempengaruhi organisasi kromatin.108 Pola metilasi DNA Aberrant yang disebabkan oleh serangan oksidatif juga mempengaruhi aktivitas perbaikan DNA.
Efek Stres Oksidatif Pada Lipid
ROS dapat menginduksi peroksidasi lipid dan mengganggu susunan lipid bilayer membran yang dapat menonaktifkan reseptor dan enzim terikat membran dan meningkatkan permeabilitas jaringan.109 Produk peroksidasi lipid, seperti MDA dan aldehida tak jenuh, mampu menginaktivasi banyak protein seluler dengan membentuk protein silang. -linkages.110-112 4-Hydroxy-2-nonenal menyebabkan penipisan GSH intraselular dan menginduksi produksi peroksida, 113,114 mengaktifkan reseptor faktor pertumbuhan epidermal, 115 dan menginduksi produksi fibronektin.116 Produk peroksidasi lipid, seperti isoprostan dan zat reaktif asam thiobarbiturat , telah digunakan sebagai biomarker tidak langsung dari stres oksidatif, dan tingkat peningkatan ditunjukkan pada kondensat napas yang dihembuskan atau cairan lavage bronchoalveolar atau paru-paru pasien penyakit paru obstruktif kronik atau perokok.117-119
Efek Stres Oksidatif pada Protein
ROS dapat menyebabkan fragmentasi rantai peptida, perubahan muatan listrik protein, penautan silang protein, dan oksidasi asam amino tertentu dan oleh karena itu menyebabkan peningkatan kerentanan terhadap proteolisis oleh degradasi oleh protease spesifik. KIMIA Sistein dan residu metionin dalam protein adalah terutama yang lebih rentan terhadap oksidasi.120 Oksidasi kelompok sulfhidril atau residu metionin dari protein menyebabkan perubahan konformasi, pembukaan protein, dan degradasi.121-8,121 Enzim yang memiliki logam pada atau dekat dengan tempat aktifnya terutama lebih sensitif terhadap oksidasi katalis logam. Modifikasi enzim oksidatif telah terbukti menghambat aktivitas mereka.123
Dalam beberapa kasus, oksidasi spesifik protein dapat terjadi. Sebagai contoh, metionin dapat dioksidasi metionin sulfoksida126 dan fenilalanin menjadi o-tirosin127; kelompok sulfhidril dapat dioksidasi untuk membentuk ikatan disulfida; gugus 128 dan karbonil dapat dimasukkan ke dalam rantai samping protein. Sinar gamma, oksidasi katalis logam, HOCl, dan ozon dapat menyebabkan pembentukan kelompok karbonil. 129
Efek Tegangan Oksidatif terhadap Transduksi Sinyal
ROS dapat menginduksi ekspresi beberapa gen yang terlibat dalam transduksi sinyal.1,130 Rasio yang tinggi untuk GSH / GSSG penting untuk melindungi sel dari kerusakan oksidatif. Gangguan rasio ini menyebabkan aktivasi faktor transkripsi sensitif redoks, seperti NF-kB, AP-1, faktor inti sel T yang diaktifkan dan faktor XIAX yang dapat diinduksi hipoksia, yang terlibat dalam respons inflamasi. Aktivasi faktor transkripsi melalui ROS dicapai dengan kaskade transduksi sinyal yang mentransmisikan informasi dari luar ke bagian dalam sel. Reseptor tirosin kinase, sebagian besar reseptor faktor pertumbuhan, seperti reseptor faktor pertumbuhan epidermal, reseptor faktor pertumbuhan endotel vaskular, dan reseptor untuk faktor pertumbuhan yang diturunkan dari trombosit, protein tirosin fosfatase, dan kinin serin / treonin adalah target ROS.1-131 Extra-seluler signal-regulated kinases, JNK, dan p133, yang merupakan anggota keluarga protein kinase mitogen yang diaktifkan dan terlibat dalam beberapa proses dalam sel termasuk proliferasi, diferensiasi, dan apoptosis, juga dapat diatur oleh oksidan.
Di bawah kondisi stres oksidatif, residu sistein di tempat pengikatan DNA c-Jun, beberapa subunit AP-1, dan kase kinase penghambat mengalami glutadriolasi reversibel. Glutaredoxin dan TRX telah dilaporkan memainkan peran penting dalam pengaturan jalur sinyal sensitif-redoks, seperti NF-kB dan AP-1, p38 mitogen-activated protein kinase, dan JNK.134-137
NF-kB dapat diaktifkan sebagai respons terhadap oksidatif kondisi stres, seperti ROS, radikal bebas, dan iradiasi UV.138 Fosforilasi IkB membebaskan NF-kB dan memungkinkannya memasuki inti untuk mengaktifkan transkripsi gen.139 Sejumlah kinase telah dilaporkan untuk memfosforilasi IkBs pada residu serin. Kinase ini adalah target dari sinyal oksidatif untuk aktivasi NF-kB.140 Reduktor meningkatkan ikatan DNA NF-kB, sedangkan oksidator menghambat pengikatan DNA NF-kB. TRX dapat melakukan tindakan yang berlawanan 2 dalam pengaturan NF-kB: di sitoplasma, menghambat degradasi IkB dan menghambat aktivasi NF-kB tetapi meningkatkan ikatan DNA NF-kB dalam nukleus.141 Aktivasi NF-kB melalui degradasi terkait oksidasi IkB menghasilkan aktivasi beberapa gen yang terkait dengan pertahanan antioksidan. NF-kB mengatur ekspresi beberapa gen yang berpartisipasi dalam respon imun, seperti IL-1b, IL-6, tumor necrosis factor-a, IL-8, dan beberapa molekul adhesi. 142,143 NF-kB juga mengatur angiogenesis dan proliferasi dan diferensiasi sel.
AP-1 juga diatur oleh negara redoks. Dengan adanya H2O2, beberapa ion logam dapat menginduksi aktivasi AP-1. Peningkatan rasio GSH / GSSG meningkatkan pengikatan AP-1 sementara GSSG menghambat pengikatan DNA pengikatan DNA AP-1.144 dari heterodimer Fos / Jun meningkat dengan pengurangan sistein tunggal yang dilestarikan dalam domain pengikat DNA masing-masing. protein, 145 sementara pengikatan DNA AP-1 dapat dihambat oleh GSSG pada banyak jenis sel, menunjukkan bahwa pembentukan ikatan disulfida oleh residu sistein menghambat pengikatan DNA AP-1.146,147 Transduksi sinyal melalui stres oksidatif dirangkum dalam Gambar 2.

KESIMPULAN
Stres oksidatif dapat timbul akibat kelebihan produksi ROS oleh reaksi metabolik yang menggunakan oksigen dan menggeser keseimbangan antara oksidan /antioksidan status yang mendukung oksidan. ROS diproduksi oleh aktivitas metabolik seluler dan faktor lingkungan, seperti polutan udara atau asap rokok. ROS adalah molekul yang sangat reaktif karena elektron yang tidak berpasangan dalam strukturnya dan bereaksi dengan beberapa makromolekul biologis dalam sel, seperti karbohidrat, asam nukleat, lipid, dan protein, dan mengubah fungsinya. ROS juga mempengaruhi ekspresi beberapa gen oleh upregulasi faktor transkripsi sensitif redoks dan pemodelan kromatin melalui perubahan asetilasi / deasetilasi histon. Peraturan negara redoks sangat penting untuk kelangsungan hidup sel, aktivasi, proliferasi, dan fungsi organ.
Referensi:
REFERENSI
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Radikal bebas, logam dan antioksidan pada kanker akibat stres oksidatif. Chem Biol Berinteraksi 2006; 160: 1-40.
2. Halliwell B, Gutteridge JMC. Radikal Bebas di Biologi dan Kedokteran. 3rd ed. New York: Oxford University Press; 1999.
3. Marnett LJ. Kerusakan peroksida lipid peroksidaseDNA oleh malondialdehida. Mutat Res. 1999; 424: 83-95.
4. Siems WG, Grune T, Esterbauer H. 4-Hidroksinonenal pembentukan selama iskemia dan reperfusi usus halus tikus. Life Sci. 1995; 57: 785–789.
5. Stadtman ER. Peran spesies oksidan dalam penuaan. Curr Med Chem. 2004; 11: 1105-1112.
6. Wang MY, Dhingra K, Hittelman WN, Liehr JG, DeAndrade M, Li DH. Lipid peroksidasi-diinduksi putatif malondialdehida-DNA adducts di jaringan payudara manusia. Kanker Epidemiol Biomarker Prev. 1996; 5: 705-710.
7. Jenner P. Stres oksidatif pada penyakit Parkinson. Ann Neurol. 2003; 53: S26-S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Penilaian kerusakan oksidatif pada protein, lipid, dan DNA di otak dari pasien dengan penyakit Alzheimer. J Neurochem. 1997; 68: 2061-2069.
9. Sayre LM, Smith MA, Perry G. Kimia dan biokimia stres oksidatif pada penyakit neurodegenerative. Curr Med Chem. 2001; 8: 721-738.
10. Toshniwal PK, Zarling EJ. Bukti untuk peningkatan peroksidasi lipid pada multiple sclerosis. Neurochem Res. 1992; 17: 205-207.
11. Dhalla NS, Temsah RM, Netticadan T. Peran stres oksidatif pada penyakit kardiovaskular. J Hipertens. 2000; 18: 655-673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, dkk. Studi tentang stres oksidatif pada model tikus hipoperfusi otak kronis. Neurochem Int. 2005; 46: 601-611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Produksi anion superoksida meningkat dalam model hipertensi genetik: peran endothelium. Hipertensi. 1999; 33: 1353-1358.
14. Kukreja RC, Hess ML. Sistem radikal bebas oksigen: dari persamaan melalui interaksi membrane protein hingga cedera kardiovaskular dan perlindungan. Cardiovasc Res. 1992; 26: 641-655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, dkk. Perokok rokok menginduksi peningkatan kerusakan DNA oksidatif, 8-hydroxydeoxyguanosine, di lokasi pusat paru-paru manusia. Karsinogenesis 1997; 18: 1763-1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Kejadian oksidatif dan nitrosatif pada asma. Radic Biol Med gratis. 2003; 35: 213-225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, dkk. Korelasi defisiensi superoksida dismutase sistemik terhadap obstruksi aliran udara pada asma. Am J Respir Crit Care Med. 2005; 172: 306-313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, dkk. Inaktivasi superoksida dismutase pada patofisiologi remodeling dan reaktivitas jalan nafas asma. Am J Pathol. 2005; 166: 663-674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Stres oksidatif dan faktor penentu di saluran pernafasan anak-anak penderita asma. Alergi. 2008; 63: 1605-1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, dkk. Stres oksidatif dan determinan genetik dan epidemiologis cedera oksidan pada asma anak. J Alergi Klinik Imunol. 2006; 118: 1097-1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Program Penelitian Asma yang parah. Airway glutathione homeostasis diubah pada anak-anak dengan asma berat: bukti untuk stres oksidan. J Alergi Klinik Imunol. 2009; 123: 146-152.
22. Miller DM, Buettner GR, Aust SD. Transisi logam sebagai katalis reaksi "autoksidasi". Radic Biol Med gratis. 1990; 8: 95-108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, Dème D, Pommier J. Mekanisme pembentukan hidrogen peroksida yang dikatalisis oleh NADPH oksidase pada membran plasma tiroid. J Biol Chem. 1991; 266: 3739-3743.
24. Granger DN. Peran xanthine oxidase dan granulocytes pada cedera ischemiareperfusion. Am J Physiol. 1988; 255: H1269-H1275.
25. Fenton HJH. Oksidasi asam tartarat dengan adanya zat besi. J Chem Soc. 1984; 65: 899-910.
26. Haber F, Weiss JJ. Dekomposisi katalitik hidrogen peroksida oleh garam besi. Proc R Soc Lond Ser A. 1934; 147: 332-351.
27. Liochev SI, Fridovich I. Haber-Weiss bersepeda beberapa tahun kemudian: sebuah pandangan alternatif. Redox Rep 70; 2002: 7-55.
28. Klebanoff SJ. Myeloperoxidase: teman dan lawan. J Leukoc Biol. 2005; 77: 598-625.
29. Whiteman M, Jenner A, Halliwell B. Modifikasi dasar yang diinduksi asam hipoklorida pada DNA timus betis yang terisolasi. Chem Res Toxicol. 1997; 10: 1240-1246.
30. Kulcharyk PA, Heinecke JW. Asam hypochlorous yang diproduksi oleh sistem myeloperoxidase dari fagosit manusia menginduksi hubungan silang kovalen antara DNA dan protein. Biokimia. 2001; 40: 3648-3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Song W, dkk. Sebuah kisah dua kontroversi: menentukan peran peroksidase dalam pembentukan nitrotyrosin secara in vivo menggunakan peroksidase eosinofil dan tikus myeloperoxidasedeficient, dan sifat spesies nitrogen reaktif peroksidase. J Biol Chem. 2002; 277: 17415-17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, et al. Degranulasi eosinofil ekstensif dan oksidasi protein perineum yang dimediasi peroksidase tidak terjadi pada model peradangan paru ovalbumin-tantangan peradangan paru. J Immunol. 2001; 167: 1672-1682.
33. van Dalen CJ, Winterbourn CC, Senthilmohan R, Kettle AJ. Nitrit sebagai substrat dan inhibitor myeloperoxidase. Implikasi untuk nitrasi dan produksi asam hypochlorous pada lokasi peradangan. J Biol Chem. 2000; 275: 11638-11644.
34. Kayu LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Peroksidasi lipid yang ditentukan oleh isoprostan plasma terkait dengan tingkat keparahan penyakit pada asma ringan. Lemak. 2000; 35: 967-974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Peningkatan 8-isoprostane, penanda stres oksidatif, pada kondensat inhalasi pasien asma. Am J Respir Crit Care Med. 1999; 160: 216-220.
36. Gereja DF, Pryor WA. Kimia radikal bebas dari asap rokok dan implikasi toksikologiknya. Perspektif Kesehatan Lingkungan. 1985; 64: 111-126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, dkk. Peradangan yang disebabkan ozon dinilai pada cairan sputum dan bronchial lavage dari penderita asma: alat noninvasif baru dalam studi epidemiologi mengenai polusi udara dan asma. Radic Biol Med gratis. 1999; 27: 1448-1454.
38. Nightingale JA, Rogers DF, Barnes PJ. Efek ozon terhirup pada oksida nitrat yang dihembuskan, fungsi paru, dan sputum yang diinduksi pada subjek normal dan penderita asma. Thorax. 1999; 54: 1061-1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, dkk. Redoks aktivitas partikulat udara di berbagai lokasi di Los Angeles Basin. Resur Lingkungan 2005; 99: 40-47.
40. Comhair SA, Thomassen MJ, Erzurum SC. Diferensial induksi glutathione peroksidase ekstraselular dan 2 nitrat oksida oksida dalam saluran udara individu sehat terpapar 100% O (2) atau asap rokok. Am J Respir Cell Mol Biol. 2000; 23: 350-354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Cedera paru yang dimediasi dengan oksidatif pada sindrom distres pernafasan akut. Crit Care Med. 1999; 27: 2028-2030.
42. Biaglow JE, Mitchell JB, Mengadakan K. Pentingnya peroksida dan superoksida dalam respons sinar-X. Int J Radiat Oncol Biol Phys. 1992; 22: 665-669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Kepekaan yang dimediasi ion tembaga terhadap situs lampiran matriks nuklir terhadap radiasi pengion. Biokimia. 1993; 32: 6214-6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Partikel Alpha menginisiasi produksi biologis anion superoksida dan hidrogen peroksida dalam sel manusia. Kanker Res. 1997; 57: 3963-3971.
45. Tuttle SW, Varnes ME, Mitchell JB, Biaglow JE. Sensitivitas terhadap oksidan kimia dan radiasi pada garis sel CHO kurang dalam aktivitas siklus pentosa oksidatif. Int J Radiat Oncol Biol Phys. 1992; 22: 671-675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, dkk. Mangan
Ekspresi gen superoksida dismutase dalam radiasi yang diinduksi
tanggapan adaptif Mol Cell Biol. 2003; 23: 2362-2378.
47. Azzam EI, Toledo de Toledo, DR Spitz, JB Kecil. Metabolisme oksidatif
memodulasi transduksi sinyal dan pembentukan mikronukleus di pengamat
sel-sel dari fibroblas manusia normal yang diiradiasi dengan iradiasi. Kanker Res.
2002; 62: 5436-5442.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Mengionisasi radiasi yang diinduksi, bergantung pada mitokondria dari reaktif
oksigen / nitrogen. Kanker Res. 2001; 61: 3894-3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, jalur Grant S. MAPK di
respon radiasi Onkogen. 2003; 22: 5885-5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, dkk. Thioredoxin
translokasi dan interaksi nuklir dengan faktor redoks-1 mengaktifkan faktor transkripsi AP-1 sebagai respons terhadap radiasi pengion. Kanker Res. 2000; 60: 6688-6695.
51. Kadet J, Douki T, Gasparutto D, Ravanat JL. Kerusakan oksidatif pada DNA: pembentukan, pengukuran dan fitur biokimia. Mutat Res. 2003; 531: 5-23.
52. Yokoya A, Cunniffe SM, O'Neill P. Efek hidrasi pada induksi istirahat untai dan lesi dasar pada film DNA plasmid dengan gammaradiasi. J Am Chem Soc. 2002; 124: 8859-8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Respon sel dan jaringan terhadap kerusakan oksidatif. Lab Invest. 1993; 69: 261-274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, dkk. Faktor nadi kappa B tergantung induksi gamma glutamylcysteine synthetase dengan mengionisasi radiasi pada sel glioblastoma manusia T98G. Radic Biol Med gratis. 1998; 24: 1256-1268.
55. Stohs SJ, Bagchi D. Mekanisme oksidatif dalam toksisitas ion logam. Radic Biol Med gratis. 1995; 18: 321-336.
56. Leonard SS, Harris GK, Shi X. Stres oksidatif yang disebabkan logam dan transduksi sinyal. Radic Biol Med gratis. 2004; 37: 1921-1942.
57. Shi H, Shi X, Liu KJ. Mekanisme oksidatif toksisitas arsenik dan karsinogenesis. Mol Cell Biochem. 2004; 255: 67-78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Mekanisme potensial untuk penurunan pembentukan oksida nitrat yang disebabkan oleh paparan oral yang berkepanjangan terhadap arsenat pada kelinci. Radic Biol Med.2003 gratis; 35: 102-113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, DNA DNA Okada S. diinduksi oleh asam dimetilarsinat, metabolit arsenik anorganik, sangat diperkuat oleh radikal anion superoksida. Biol Pharm Bull. 1995; 18: 45-58.
60. Anggota parlemen Waalkes, Liu J, Ward JM, Diwan LA. Mekanisme yang mendasari karsinogenesis arsenik: hipersensitifitas tikus yang terpapar arsen anorganik selama masa gestasi. Toksikologi. 2004; 198: 31-38.
61. Schiller CM, Fowler BA, Woods JS. Efek arsen pada aktivasi piruvat dehidrogenase. Perspektif Kesehatan Lingkungan. 1977; 19: 205-207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Keterlibatan radikal bebas pada porfiria neurologis dan keracunan timah. Mol Cell Biochem. 1991; 103: 73-83.
63. Tripathi RM, Raghunath R, Mahapatra S. Timbal darah dan pengaruhnya pada kadar Cd, Cu, Zn, Fe dan hemoglobin anak-anak. Sci Total Lingkungan. 2001; 277: 161-168.
64. Nehru B, Dua R. Efek selenium diet pada timbal neurotoksisitas. J Lingkungan Patol Toxicol Oncol. 1997; 16: 47-50.
65. Reid TM, Feig DI, Loeb LA. Mutagenesis oleh radikal oksigen yang disebabkan logam. Perspektif Kesehatan Lingkungan. 1994; 102 (suppl 3): 57-61.
66. Kinnula VL, Crapo JD. Superoksida dismutases di paru-paru dan penyakit paru-paru manusia. Am J Respir Crit Care Med. 2003; 167: 1600-1619.
67. Kinnula VL. Produksi dan degradasi metabolit oksigen selama keadaan inflamasi di paru-paru manusia. Curr Drug Targetkan Alergi Inflamasi. 2005; 4: 465-470.
68. Zelko IN, Mariani TJ, Folz RJ. Keluarga superoksida dismutase multigene: perbandingan struktur gen, evolusi, dan ekspresi CuZn-SOD (SOD1), Mn-SOD (SOD2), dan EC-SOD (SOD3); Radic Biol Med gratis. 2002; 33: 337-349.
69. Kirkman HN, Rolfo M, Ferrari AM, Gaetani GF. Mekanisme proteksi katalase oleh NADPH. Kinetika dan stoikiometri. J Biol Chem. 1999; 274: 13908-13914.
70. Flohé L. Glutathione peroksidase. Basic Life Sci. 1988; 49: 663-668.
71. Arthur JR. Glutathione peroksidase. Cell Mol Life Sci. 2000; 57: 1825-1835.
72. Chu FF, Doroshow JH, Esworthy RS. Ekspresi, karakterisasi, dan distribusi jaringan peroksidase glutathione selenium yang bergantung pada selenium, GSHPx-GI. J Biol Chem. 1993; 268: 2571-2576.
73. Comhair SA, PR Bhathena, Farver C, Thunnissen FB, Erzurum SC. Induksi peroksidase glutathione ekstraseluler pada paru-paru asma: bukti regulasi redoks ekspresi pada sel epitel jalan nafas manusia. FASEB J. 2001; 15: 70-78.
74. Gromer S, Urig S, Becker K. Sistem thioredoxin dari sains ke klinik. Med Res Rev. 2004; 24: 40-89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, Pääkkö P, dkk. Ekspresi spesifik sel peroksiredoksin pada paru-paru manusia dan sarkoidosis paru. Thorax. 2002; 57: 157-164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, dkk. Manusia peroxiredoxin 5 adalah peroxynitrite reductase. FEBS Lett. 2004; 571: 161-165.
77. Holmgren A. Fungsi antioksidan sistem thioredoxin dan glutaredoxin. Antioksidan Redoks Sinyal. 2000; 2: 811-820.
78. Dickinson DA, Forman HJ. Glutathione dalam pertahanan dan pensinyalan: pelajaran dari tiol kecil. Ann NY Acad Sci. 2002; 973: 488-504.
79. Sies H. Glutathione dan perannya dalam fungsi seluler. Radic Biol Med gratis. 1999; 27: 916-921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Jalur evolusioner paralel untuk transferase glutathione: struktur dan mekanisme enzim kappa kelas mitokondria rGSTK1-1. Biokimia. 2004; 43: 52-61.
81. Robinson A, Huttley GA, Booth HS, Papan PG. Pemodelan dan studi bioinformatika kelas kappa manusia glutathione transferase memprediksi keluarga transferase ketiga yang baru dengan homologi menjadi isomerase 2-hydroxychromene-2-carboxylate prokariotik. Biochem J. 2004; 379: 541-552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Gambaran struktural umum dari MAPEGda melebarkan sayap luas protein membran terkait dengan fungsi yang sangat berbeda dalam metabolisme eicosanoid dan glutathione. Protein Sci. 1999; 8: 689-692.
83. Hayes JD, Pulford DJ. Keluarga supergen glutathione S-transferase: regulasi GST dan kontribusi isoenzim terhadap kemoproteksi kanker dan resistansi obat. Crit Rev Biochem Mol Biol. 1995; 30: 445-600.
84. Armstrong RN. Struktur, mekanisme katalitik, dan evolusi pemindahan glutathione. Chem Res Toxicol. 1997; 10: 2-18.
85. Hayes JD, McLellan LI. Glutathione dan enzim yang bergantung glutathione mewakili pertahanan yang diatur secara ordinat melawan stres oksidatif. Radic Res. 1999; 31: 273-300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Struktur, fungsi dan evolusi transferase glutathione: implikasi untuk klasifikasi anggota nonmamalia superfamili enzim purba. Biochem J. 2001; 360: 1-16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, dkk. Glutathione S-transferase Mu memodulasi sinyal stres yang diaktifkan dengan menekan apoptosis sinyal-mengatur kinase 1. J Biol Chem. 2001; 276: 12749-12755.
88. Dorion S, Lambert H, Landry J. Aktivasi jalur sinyal p38 oleh sengatan panas melibatkan disosiasi glutathione S-transferase Mu dari Ask1. J Biol Chem. 2002; 277: 30792-30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L, dkk. Peraturan pensinyalan JNK oleh GSTp. EMBO J. 1999; 18: 1321-1334.
90. Manevich Y, Feinstein SI, Fisher AB. Aktivasi enzim antioksidan 1-CYS peroxiredoxin memerlukan glutathionylation yang dimediasi oleh heterodimerisasi dengan pGST. Proc Natl Acad Sci US A. 2004; 101: 3780-3785.
91. Bunker VW. Radikal bebas, antioksidan dan penuaan. Med Lab Sci. 1992; 49: 299-312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, et al. Stres oksidatif sistemik dan hubungannya dengan usia dan penyakit. J Am Geriatr Soc. 1996; 44: 823-827.
93. White E, Shannon JS, Patterson RE. Hubungan antara vitamin dan
penggunaan suplemen kalsium dan kanker usus besar. Kanker Epidemiol Biomarker Prev. 1997; 6: 769-774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Mekanisme Novel dari senyawa antioksidan alami dalam sistem biologi: keterlibatan enzim glutathione dan glutathione. J Nutr Biochem. 2005; 16: 577-586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Perubahan status glutathione jantung setelah iskemia dan reperfusi. Experientia. 1985; 41: 42-43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. Kimia radikal karotenoid dan sifat antioksidan / pro-oksidan. Arch Biochem Biofis. 2004; 430: 37-48.
97. Rice-Evans CA, Sampson J, Bramley PM, Holloway DE. Mengapa kita mengharapkan karotenoid menjadi antioksidan in vivo? Radic Res. 1997; 26: 381-398.
98. Niles RM. Jalur pensinyalan dalam kemoprevensi retinoid dan pengobatan kanker. Mutat Res. 2004; 555: 81-96.
99. Donato LJ, Noy N. Penekanan pada pertumbuhan karsinoma mammae oleh asam retinoat: gen proapoptotik menjadi target reseptor asam retinoat dan pensinyalan protein pengikat asam amino retinoat seluler. Kanker Res. 2005; 65: 8193-8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, dkk. Bcl-2 adalah pengatur utama untuk kematian sel apoptotik akibat asam retinoat dalam neuroblastoma. Onkogen. 2006; 25: 5046-5055.
101. Dalton TP, Shertzer HG, Puga A. Peraturan ekspresi gen oleh oksigen reaktif. Ann Rev Pharmacol Toxicol. 1999; 39: 67-101.
102. Scandalios JG. Respon genom terhadap stres oksidatif. In: Meyers RA, ed. Ensiklopedia Biologi Sel Molekuler dan Pengobatan Molekuler. Vol 5 2nd ed. Weinheim, Jerman: Wiley-VCH; 2004: 489-512.
103. Ghosh R, Mitchell DL. Efek kerusakan DNA oksidatif pada elemen promotor pada faktor transkripsi mengikat. Asam Nukleat Res. 1999; 27: 3213-3218.
104. Marietta C, Gulam H, Brooks PJ. Lesi 8, 50-cyclo-20-deoxyadenosine tunggal dalam kotak TATA mencegah pengikatan protein pengikat TATA dan sangat mengurangi transkripsi secara in vivo. Perbaikan DNA (Amst). 2002; 1: 967-975.
105. Jackson AL, Chen R, Loeb LA. Induksi ketidakstabilan mikrosatelit
dengan kerusakan DNA oksidatif. Proc Natl Acad Sci US A. 1998; 95: 12468-12473.
106. Caldecott KW. Interaksi protein protein selama perbaikan DNA untai tunggal mamalia. Biochem Soc Trans. 2003; 31: 247-251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Kerusakan DNA oksidatif: mekanisme, mutasi, dan penyakit. FASEB J. 2003; 17: 1195-1214.
108. Jones PL, Wolffe AP. Hubungan antara organisasi kromatin dan metilasi DNA dalam menentukan ekspresi gen. Semin Cancer Biol. 1999; 9: 339-347.
109. Girotti AW. Mekanisme peroksidasi lipid. J Free Radic Biol Med. 1985; 1: 87-95.
110. Siu GM, Draper HH. Metabolisme malonaldehida in vivo dan in vitro. Lemak. 1982; 17: 349-355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. Kemungkinan keterlibatan produk peroksidasi lipid 4-hidroksinonenal dalam pembentukan kromolipid neon. Biochem J. 1986; 239: 405-409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Perubahan tergantung usia pada tingkat peroksida lipid dalam fraksi lipoprotein serum manusia. J Gerontol. 1984; 39: 269-272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, dkk. 4- Hydroxynonenal, produk aldehid dari peroksidasi lipid membran, mengganggu transport glutamat dan fungsi mitokondria di synaptosom. Neuroscience. 1997; 806: 85-96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Aktivasi jalur pensinyalan stres oleh produk akhir peroksidasi lipid. 4-hydroxy-2-nonenal adalah induser potensial dari produksi peroksida intraselular. J Biol Chem. 1999; 274: 2234-2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Aktivasi reseptor EGF oleh LDL teroksidasi. FASEB J. 1998; 12: 665-671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal meningkatkan produksi fibronektin oleh fibroblok paru manusia IMR-90 sebagian melalui aktivasi protein fase-reseptor yang terkait reseptor ekstraselular reseptor kinase jalur p44 / 42. Toxicol Appl Pharmacol. 2002; 184: 127-135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. Menghirup 8-isoprostane sebagai biomarker in vivo pada tekanan oksidatif paru pada pasien PPOK dan perokok sehat. Am J Respir Crit Care Med. 2000; 162: 1175-1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Ketahanan permeabilitas, peradangan, dan stres oksidan di ruang udara perokok. Am J Respir Crit Care Med. 1999; 159: 473-479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Peningkatan kandungan zat reaktif asam thiobarbiturat dan hidrogen peroksida pada kondensat napas kadaluarsa pasien dengan penyakit paru obstruktif kronik yang stabil: tidak ada efek merokok yang signifikan. Respir Med. 1999; 93: 389-396.
120. Kelly FJ, Mudway IS. Oksidasi protein pada antarmuka udara-paru. Asam amino. 2003; 25: 375-396.
121. Dean RT, Roberts CR, Jessup W. Fragmentasi polipeptida ekstraselular dan intraseluler oleh radikal bebas. Prog Clin Biol Res. 1985; 180: 341-350.
122. Keck RG. Penggunaan t-butyl hydroperoxide sebagai probe untuk oksidasi metionin dalam protein. Anal Biochem. 1996; 236: 56-62.
123. Davies KJ. Kerusakan dan degradasi protein oleh radikal oksigen. I. Aspek umum. J Biol Chem. 1987; 262: 9895-9901.
124. Stadtman ER. Oksidasi katalis ion logam dari protein: mekanisme biokimia dan konsekuensi biologis. Radic Biol Med gratis.
1990; 9: 315-325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Inaktivasi enzim metabolik utama oleh reaksi oksidasi fungsi campuran: kemungkinan implikasi dalam perputaran protein dan penuaan. Proc Natl Acad Sci US A. 1983; 80: 1521-1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oksidasi residu metionin protein: konsekuensi biologis. Antioksidan Redoks Sinyal. 2003; 5: 577-582.
127. Stadtman ER, Levine RL. Oksidasi bebas radikal bebas dari asam amino bebas dan residu asam amino dalam protein. Asam amino. 2003; 25: 207-218.
128. Stadtman ER. Oksidasi protein dalam penuaan dan penyakit terkait usia. Ann NY Acad Sci. 2001; 928: 22-38.
129. Shacter E. Kuantifikasi dan signifikansi oksidasi protein pada sampel biologis. Obat Metab Rev. 2000; 32: 307-326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Tegangan oksidatif dan pensinyalan sel. Curr Med Chem. 2004; 11: 1163-1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Faktor pertumbuhan endotel vaskular (VEGF) dan reseptornya. FASEB J. 1999; 13: 9-22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, dkk. Regulasi generasi spesies reaktif-oksigen dalam fibroblas oleh Rac1. Biochem J. 1996; 318: 379-382.
133. Sun T, Oberley LW. Peraturan Redoks aktivator transkripsi. Radic Biol Med gratis. 1996; 21: 335-348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redox pengaturan DNA c-Jun yang diikat dengan glutokhiolasi reversibel. FASEB J. 1999; 13: 1481-1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. Deteksi situ protein S-glutathionylated mengikuti derivatisasi sistein yang dikatalisis glutaredoksin-1. Biochim Biofis Acta. 2006; 1760: 380-387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulasi glutaredoxin-1
Ekspresi dalam model tikus penyakit saluran napas alergi. Am J Respir Cell Mol Biol. 2007; 36: 147-151.
137. Filomeni G, Rotilio G, Ciriolo MR. Cell signaling dan sistem redoks glutathione. Biochem Pharmacol. 2002; 64: 1057-1064.
138. Pande V, Ramos MJ. Pengenalan molekuler 15-deoxydelta (12,14) prostaglandin J (2) oleh faktor-kappa B dan protein seluler lainnya. Bioorg Med Chem Lett. 2005; 15: 4057-4063.
139. Perkins ND. Mengintegrasikan jalur pensinyalan sel dengan fungsi NF-kappaB dan IKK. Nat Rev Mol Cell Biol. 2007; 8: 49-62.
140. Gilmore TD. Pengantar NF-kappaB: pemain, jalur, perspektif. Onkogen. 2006; 25: 6680-6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Perbedaan peran thioredoxin di sitoplasma dan di nukleus. Mekanisme dua langkah regulasi redoks faktor transkripsi NF-kappaB. J Biol Chem. 1999; 274: 27891-27897.
142. Ward PA. Peran pelengkap, kemokin dan sitokin regulasi pada cedera paru akut. Ann NY Acad Sci. 1996; 796: 104-112.
143. Akira S, Kishimoto A. NF-IL6 dan NF-kB dalam regulasi gen sitokin. Adv Immunol. 1997; 65: 1-46.
144. Meyer M, Schreck R, Baeuerle PA. H2O2 dan antioksidan memiliki efek berlawanan pada aktivasi NF-kappa B dan AP-1 pada sel utuh: AP-1 sebagai faktor responsif antioksidan sekunder. EMBO J. 1993; 12: 2005-2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Redox mengatur aktivitas pengikatan DNA fos dan jun secara in vitro. Ilmu. 1990; 249: 1157-1161.
146. Galter D, Mihm S, Droge W. Efek berbeda dari disulfida glutathione pada faktor transkripsi nuklir kB dan protein penggerak-1. Eur J Biochem. 1994; 221: 639-648.
147. Hirarki K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. AP-1 Aktivitas transkripsi diatur oleh hubungan langsung antara thioredoxin dan Ref-1. Proc Natl Acad Sci US A. 1997; 94: 3633-3638.
Posting Penafian
Informasi di sini tentang "El Paso, TX Stres Oksidatif dan Antioksidan" tidak dimaksudkan untuk menggantikan hubungan pribadi dengan profesional perawatan kesehatan yang berkualifikasi atau dokter berlisensi dan bukan merupakan saran medis. Kami mendorong Anda untuk membuat keputusan perawatan kesehatan berdasarkan penelitian dan kemitraan Anda dengan profesional perawatan kesehatan yang berkualifikasi.
Informasi Blog & Ruang Lingkup Diskusi
Lingkup informasi kami terbatas pada Chiropraktik, muskuloskeletal, akupunktur, obat-obatan fisik, kesehatan, berkontribusi etiologis gangguan viscerosoma dalam presentasi klinis, dinamika klinis refleks somatovisceral terkait, kompleks subluksasi, masalah kesehatan sensitif, dan/atau artikel, topik, dan diskusi kedokteran fungsional.
Kami menyediakan dan menyajikan kerjasama klinis dengan para ahli dari berbagai disiplin ilmu. Setiap spesialis diatur oleh ruang lingkup praktik profesional mereka dan yurisdiksi lisensi mereka. Kami menggunakan protokol kesehatan & kebugaran fungsional untuk merawat dan mendukung perawatan cedera atau gangguan pada sistem muskuloskeletal.
Video, postingan, topik, subjek, dan wawasan kami mencakup masalah, masalah, dan topik klinis yang terkait dengan dan secara langsung atau tidak langsung mendukung ruang lingkup praktik klinis kami.*
Kantor kami telah berupaya memberikan kutipan yang mendukung dan telah mengidentifikasi studi penelitian relevan yang mendukung postingan kami. Kami menyediakan salinan studi penelitian pendukung yang tersedia untuk dewan pengawas dan publik atas permintaan.
Kami memahami bahwa kami mencakup hal-hal yang memerlukan penjelasan tambahan tentang bagaimana hal itu dapat membantu dalam rencana perawatan atau protokol perawatan tertentu; oleh karena itu, untuk membahas lebih lanjut materi pelajaran di atas, jangan ragu untuk bertanya Dr Alex Jimenez, DC, atau hubungi kami di 915-850-0900.
Kami di sini untuk membantu Anda dan keluarga Anda.
Berkah
Dr. Alex Jimenez IKLAN, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
email: pelatih@elpasofungsionalmedicine.com
Lisensi sebagai Doctor of Chiropractic (DC) di Texas & New Mexico*
Lisensi Texas DC # TX5807, Lisensi New Mexico DC # NM-DC2182
Lisensi sebagai Perawat Terdaftar (RN*) di Florida
Lisensi Florida Lisensi RN # RN9617241 (Kontrol No. 3558029)
Status Kompak: Lisensi Multi-Negara: Berwenang untuk Praktek di Status 40*
Saat ini Matrikulasi: ICHS: MSN* FNP (Program Praktisi Perawat Keluarga)
Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Kartu Bisnis Digital Saya
 Sekali lagi Kami Menyambut Anda¸
Sekali lagi Kami Menyambut Anda¸
